



巢鼠属 (MicromysDehne, 1841) 隶属于啮齿目 (Rodentia) 鼠科 (Muridae),是最小的啮齿动物之一,体重一般10 g左右,体长40 ~ 80 mm。巢鼠体小灵活,喜欢攀爬,常在植物穗和枝条间攀援觅食,善于用植物茎、叶等编制球形小巢,悬挂于离地面 100 cm 左右的植物茎秆或树枝上 (Smith 和解焱,2009)。它们以植物性食物为主,包括野生植物和多种农作物的种子、根和绿色部分,有时也食虫 (盛和林,2005)。该属在亚洲和欧洲广泛分布 (Abramov et al., 2009; Råberg et al., 2013),目前该属共分为两个物种:巢鼠 (M. minutus Pallas, 1771) 和红耳巢鼠 (M. erythrotisBlyth, 1856)。但由于缺少系统的研究,巢鼠属的分类和分布仍存在争议。
巢鼠长期以来被认为是该属的唯一物种,在亚洲和欧洲广泛分布(Wilson and Reeder, 2005; Abramov et al., 2009),分布区包括法国、芬兰、挪威、澳大利亚、德国、意大利、瑞典、俄罗斯、中国、印度、日本、韩国、缅甸等多个国家 (Abramov et al., 2009; Råberg et al., 2013)。在中国,巢鼠广泛分布于除沙漠、荒漠和青藏高原高海拔以外的区域 (刘少英和吴毅,2019)。王应祥 (2003) 根据分布地区和形态差异将中国的巢鼠划分出7个亚种和3个居群。
红耳巢鼠最早由Blyth (1856) 基于印度阿萨姆卡西山的标本进行描记,此后长期被作为巢鼠的一个亚种 (Wilson and Reeder, 2005)。Yasuda等 (2005) 结合Cyt b、COI、IRBP 基因分析认为成都标本可能是一个不同于巢鼠的独立物种。Abramov 等 (2009) 通过形态和遗传分析认为中国成都和越南北部的巢鼠应为一个独立物种,遂将红耳巢鼠提升为有效种。但红耳巢鼠的分类地位目前仍存在争议。虽然一些学者承认其有效种地位(蒋志刚等,2015),但由于标本数量较少,这种分类体系并未被最新的《世界哺乳动物手册》(Handbook of the Mammals of the World) 所采纳 (Wilson and Mittermeier, 2018)。目前已知红耳巢鼠分布于中国、越南、缅甸和印度,在中国其主要分布于西藏、云南、四川等西南地区,此外福建有一个间断分布的种群 (Abramov et al., 2009;蒋志刚等,2015)。
2019年11月,在安徽省清凉峰 (北纬30°00′, 东经118°10′) 开展小型兽类调查时采集到8号巢鼠属标本,形态对比和分子系统发育分析显示其包含巢鼠 (采集号:QLF1911124、QLF1911125、QLF1911161和QLF1911162) 和红耳巢鼠 (采集号:QLF1911037、QLF1911163、QLF1911183和QLF1911340) 两个物种,支持红耳巢鼠的有效种地位。基于这两种标本的形态差异,本文结合现有文献资料和国家动物标本资源库标本照片对中国巢鼠属的分布进行了整理。
1 研究方法
1.1 形态数据测量和分析
本研究在安徽清凉峰海拔1 000 ~ 1 700 m的草丛中采集到8号巢鼠属标本,所有标本的皮张、头骨和肌肉组织均保藏于安徽师范大学动物标本馆。首先将这8号巢鼠的外形和头骨标本与彭鸿绶和王应祥 (1981) 以及 Abramov 等 (2009) 对巢鼠和红耳巢鼠的描述进行对比。然后依据夏霖等 (2006) 和 Abramov 等 (2009) 选取并测量了5个外形指标和11个头骨指标:体重 (Body weight, W)、头体长 (Head and body length, HB)、尾长 (Tail length, TL)、后足长 (Hind foot length, HF)、耳长 (Ear length, EL)、颅全长 (Greatest length of skull, GLS)、眶间宽 (Interorbital breadth, IB)、颅宽 (Braincase breadth, BB)、颧宽 (Zygomatic width, ZW)、颅高 (Braincase height, BH)、臼齿横宽 (Breadth across upper molars, BUM)、门齿孔长 (Length of foramina incisive, FIL)、髁基长 (Condylobasal length, CL)、腭长 (Palatal length, PL)、上臼齿长 (Upper molar row length, UML) 和下臼齿长 (Lower molar row length, LML)。
1.2 遗传分析
利用DNA提取试剂盒提取这8号标本的总DNA,通过PCR扩增其Cyt b基因送至安徽通用生物系统有限公司进行测序。测序返回后,使用DNASTAR软件的子程序Seqman进行序列校准拼接。通过NCBI网站进行Blast比对得到同源序列,并从GenBank现有91条巢鼠Cyt b序列中下载了22条来自中国 (四川、台湾)、越南、韩国、日本、俄罗斯、瑞士、法国和芬兰的巢鼠属物种的Cyt b序列,使用 MEGA-X (Kumar et al., 2018) 进行序列比对,并基于Kimura 2-parameter进行遗传距离分析。利用Phylosuite (Zhang et al., 2020) 构建最大似然树 (maximum likelihood)。
1.3 中国巢鼠属分布分析
基于巢鼠与红耳巢鼠的形态差异特征,我们对国家动物标本资源库 (
2 结果
2.1 形态比较
本次采集的8号标本均为成体,依据外形分为两种。其中4号 (QLF1911124 雄性、QLF1911125 雄性、QLF1911161 雌性和 QLF1911162 雄性) 背毛棕红色,毛基深灰色,体侧近腹面呈棕黄色;腹部毛基灰色,毛尖为白色。耳内外均被毛,耳后缘具三角形耳屏,耳瓣毛簇淡白色到浅黄色。尾背面褐色,腹面土黄色,前后足皆为浅黄色 (图1)。头骨狭小,脑颅略有隆起 (图2)。这些特点与巢鼠的描记一致 (盛和林,2005;Smith 和解焱,2009)。另外4号标本 (QLF1911037 雄性、QLF1911163雄性、QLF1911183雌性和 QLF1911340 雌性) 背毛灰色,毛尖棕色;腹部毛基黄色,毛尖灰色,背腹异色不明显。耳内外均被毛,耳后缘具三角形耳屏,耳瓣毛簇黑红色。尾明显大于头体长 (TL/HB ≈ 110 %),尾黑褐色,上下近乎同色,前足和后足背都为灰色 (图1)。与巢鼠相比,其头骨隆起更明显 (图2)。这些特征与对红耳巢鼠的描记一致,但体型偏小 (Abramov et al., 2009) (表1)。
图1
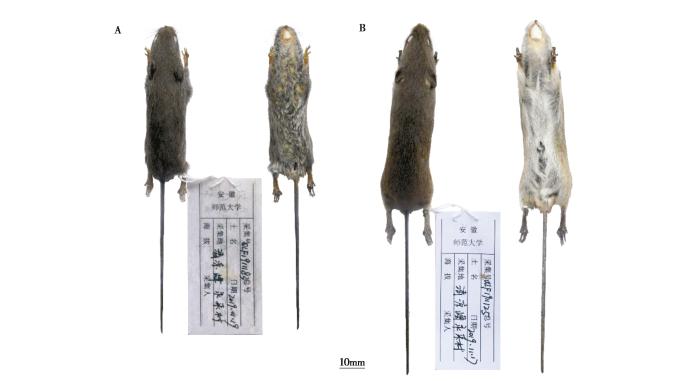
图1
安徽清凉峰红耳巢鼠 (A:QLF1911183,雌性) 和巢鼠 (B:QLF1911125,雄性) 皮张标本
Fig. 1
The skins of M. erythrotis (A: QLF1911183, female) and M. minutus (B: QLF1911125, male) from Qingliangfeng in Anhui Province
图2
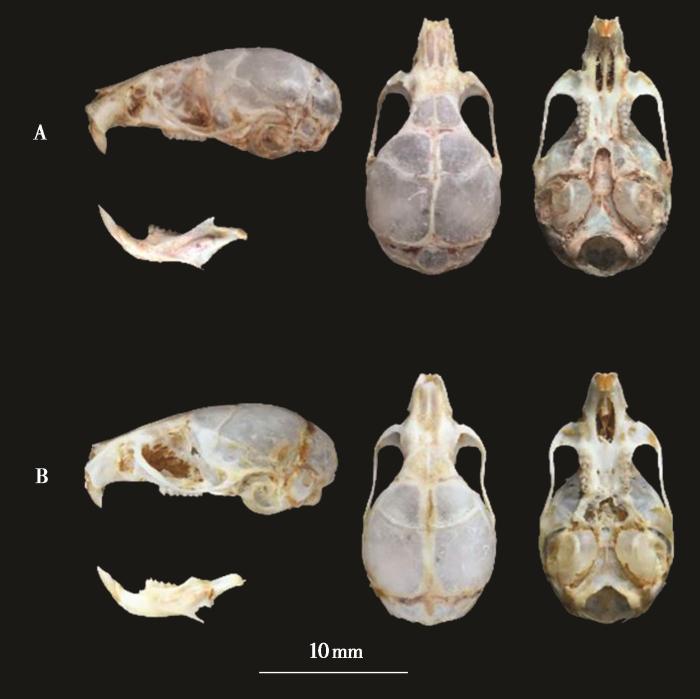
图2
安徽清凉峰红耳巢鼠(A:QLF1911183,雌性)和巢鼠(B:QLF1911125,雄性)头骨
Fig. 2
Cranium of M. erythrotis (A: QLF1911183, female) and M.minutus (B: QLF1911125, male) from Qingliangfeng in Anhui Province
表1 安徽巢鼠属标本的外形、头骨量度及其与Abramov等 (2009) 的对比
Table 1
| 测量指标 Measurements | 红耳巢鼠 M. erythrotis | 巢鼠 M. minutus | |||||
|---|---|---|---|---|---|---|---|
| 中国安徽 | 越南 | 中国四川 | 中国安徽 | 欧洲 | 俄罗斯 | ||
| Anhui, China | Vietnam | Sichuan, China | Anhui,China | Europe | Russia | ||
| n = 4 | n = 8 | n = 9 | n = 4 | n= 44 | n = 14 | ||
| 体重Body weight | 6.54 ± 0.68 | N.A. | N.A. | 6.80 ± 0.35 | N.A. | N.A. | |
| 头体长Head and body length | 55.75 ± 3.27 | 61.11 ± 7.58 | N.A. | 55.00 ± 1.87 | N.A. | N.A. | |
| 尾长Tail length | 61.75 ± 2.59 | 75.33 ± 11.17 | N.A. | 66.50 ± 2.50 | N.A. | N.A. | |
| 后足长Hind foot length | 14.00 ± 0.71 | 15.13 ± 1.62 | N.A. | 14.87 ± 0.22 | N.A. | N.A. | |
| 耳长Ear length | 9.63 ± 0.41 | 10.67 ± 0.94 | N.A. | 10.00 ± 0.00 | N.A. | N.A. | |
| 颅全长Greatest length of skull | 17.36 ± 0.09 | 18.45 ± 0.46 | 18.65 ± 0.65 | 17.45 ± 0.51 | 17.08 ± 0.39 | 17.02 ± 0.87 | |
| 髁基长Condylobasal length | 16.35 ± 0.19 | 17.12 ± 0.65 | 17.07 ± 0.76 | 16.32 ± 0.19 | 15.85 ± 0.37 | 15.99 ± 0.80 | |
| 腭长Palatal length | 8.58 ± 0.10 | 9.20 ± 0.23 | 9.25 ± 0.25 | 8.79 ± 0.22 | 8.56 ± 0.22 | 8.57 ± 0.50 | |
| 眶间宽Interorbital breadth | 3.20 ± 0.04 | 3.31 ± 0.17 | 3.22 ± 0.17 | 3.21 ± 0.07 | 3.21 ± 0.08 | 3.18 ± 0.12 | |
| 颅宽Braincase breadth | 9.07 ± 0.03 | 8.96 ± 0.30 | 9.31 ± 0.21 | 9.19 ± 0.07 | 8.68 ± 0.28 | 8.68 ± 0.16 | |
| 颅高Braincase height | 7.14 ± 0.21 | 7.05 ± 0.29 | 7.12 ± 0.38 | 7.03 ± 0.11 | 6.76 ± 0.25 | 6.80 ± 0.16 | |
| 颧宽Zygomatic width | 9.12 ± 0.02 | 8.78 ± 0.48 | 9.27 ± 0.20 | 9.27 ± 0.13 | 8.88 ± 0.24 | 8.79 ± 0.34 | |
| 上臼齿长Upper molar row length | 2.83 ± 0.02 | 2.84 ± 0.07 | 2.98 ± 0.10 | 2.99 ± 0.07 | 2.73 ± 0.09 | 2.74 ± 0.12 | |
| 下臼齿长Lower molar row length | 2.79 ± 0.03 | 2.77 ± 0.04 | 2.88 ± 0.07 | 2.88 ± 0.07 | 2.69 ± 0.10 | 2.69 ± 0.08 | |
| 臼齿横宽Breadth across upper molars | 3.57 ± 0.02 | 3.58 ± 0.19 | 3.44 ± 0.26 | 3.70 ± 0.07 | 2.91 ± 0.22 | 2.69 ± 0.21 | |
| 门齿孔长Length of foramina incisive | 3.51 ± 0.03 | 3.50 ± 0.19 | 3.63 ± 0.26 | 3.63 ± 0.09 | 3.52 ± 0.08 | 3.48 ± 0.12 | |
N.A.:无数据 No data
2.2 系统发育分析
这8号巢鼠标本的Cyt b序列全长 1 140 bp,并提交GenBank (登陆号:MW585766, MW585780, MW585804 ~ MW585809)。基于最大似然法构建的Cyt b 系统发育树显示这8号标本分为两个明显的分支:其中4号与巢鼠聚为一支,另外4号与红耳巢鼠聚为一支 (BS = 100 %; 图3);两种巢鼠间的遗传距离达13.3% (表2)。这些结果都支持红耳巢鼠作为一个种。基于Kimura 2-parameter 模型估算显示安徽巢鼠与俄罗斯等地巢鼠的遗传距离为 0.97% ~ 1.06%;安徽红耳巢鼠与越南标本的遗传距离为0.88%,与四川标本的遗传距离为3.35% (表2)。国内不同区域红耳巢鼠存在遗传分化,安徽红耳巢鼠标本与越南标本亲缘关系更为接近。系统发育分析显示巢鼠可分为3个支系:俄罗斯、中国台湾和安徽标本分为一支;日本、韩国、欧洲标本分为一支;两条产地为中国的序列 (HM217360和HM217361) 单独聚为一支 (图3)。
图3
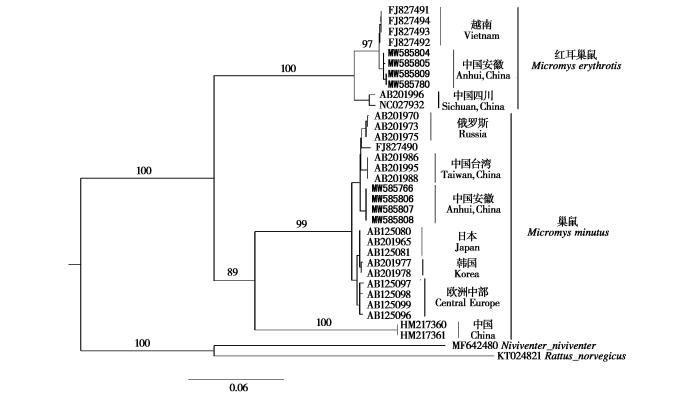
图3
基于线粒体基因Cyt b构建的最大似然树
Fig.3
The maximum likelihood tree based on mitochondrial Cyt b gene
表2 巢鼠属线粒体基因Cyt b遗传距离
Table 2
物种(地区) Species (Region) | 红耳巢鼠 (中国安徽) M. erythrotis (Anhui,China) | 红耳巢鼠 (越南) M. erythrotis (Vietnam) | 红耳巢鼠 (中国四川) M. erythrotis (Sichuan,China) | 巢鼠 (中国安徽) M. minutus (Anhui,China) | 巢鼠 (俄罗斯) M. minutus (Russia) | 巢鼠 (中国台湾) M. minutus (Taiwan,China) |
|---|---|---|---|---|---|---|
红耳巢鼠(中国安徽) M. erythrotis (Anhui,China) | *** | |||||
红耳巢鼠(越南) M. erythrotis (Vietnam) | 0.0088 | *** | ||||
红耳巢鼠(中国四川) M. erythrotis (Sichuan,China) | 0.0335 | 0.0298 | *** | |||
巢鼠(中国安徽) M. minutus (Anhui,China) | 0.1333 | 0.1300 | 0.1279 | *** | ||
巢鼠(俄罗斯) M. minutus (Russia) | 0.1390 | 0.1356 | 0.1291 | 0.0106 | *** | |
巢鼠(中国台湾) M. minutus (Taiwan,China) | 0.1346 | 0.1335 | 0.1291 | 0.0097 | 0.0097 | *** |
2.3 巢鼠属物种在中国的分布
我们从国家动物标本资源库共下载了231张有分类学特征、可用于鉴定的巢鼠属标本照片,这些照片共包含77号巢鼠标本。我们基于上述巢鼠与红耳巢鼠的形态差异对这些标本进行了鉴定,其中有9号为巢鼠,分布于陕西和黑龙江;68号为红耳巢鼠,分布于云南、四川、陕西、湖北、西藏、贵州、重庆,两个物种在安徽清凉峰同域分布,在陕西镇巴县、城固县都有记录,可能为同域分布 (图4,附表1)。此外根据文献记载,除上述区域外,巢鼠在我国吉林、辽宁、内蒙古、河北、甘肃、新疆、江苏、安徽、浙江、湖南、江西、广东、福建、台湾也有记录;红耳巢鼠在福建和广西也有分布 (Abramov et al., 2009;蒋志刚等,2015)。但由于红耳巢鼠长期作为巢鼠的一个亚种,这些巢鼠的记录中可能会包含有红耳巢鼠,对已有巢鼠标本的进一步整理,可能会发现更多红耳巢鼠的分布区。
图4

3 讨论
长期以来,巢鼠属被认为是一个单型属,只包含巢鼠一个物种 (Wilson and Reeder, 2005)。Abramov等 ( 2009) 认为红耳巢鼠应为一个独立物种,但因为数据较少,并没有被主流的分类体系所承认 (Wilson and Mittermeier, 2018)。本研究在安徽清凉峰海拔1 000 ~ 1 700 m的草丛中采集到8号巢鼠属标本。通过分子、外形和头骨特征,证明这些标本包含巢鼠和红耳巢鼠两个物种。在安徽清凉峰,红耳巢鼠在海拔 1 000 ~ 1 700 m区间皆有分布,巢鼠仅分布在海拔1 600 m,两种巢鼠在海拔1 600 m同域分布,支持红耳巢鼠为一独立物种。
已有研究结果显示红耳巢鼠主要分布于越南北部,印度阿萨姆和我国西藏、云南、四川等中国西南地区,但在福建有一个间断分布的种群,这两个分布区被巢鼠所隔断 (Abramov et al., 2009;蒋志刚等,2015)。安徽介于红耳巢鼠中国西部地区和福建分布区之间,填补了该物种的分布空缺。通过整理国家动物标本资源库的巢鼠属标本,发现红耳巢鼠在云南、四川、陕西、湖北、西藏、贵州、重庆都有分布,除安徽清凉峰外,红耳巢鼠和巢鼠在陕西镇巴县和城固县也都有记录。基于已有的数据显示红耳巢鼠的分布区仍存在明显的间断,在其两个分布区之间的湖南、江西、广东目前只有巢鼠分布纪录。鉴于红耳巢鼠长期作为巢鼠的一个亚种,我们推测该地区的“巢鼠”中可能混杂着红耳巢鼠。获取更多该区域标本,对中国巢鼠属的分布研究具有重要意义。
通过整合GenBank中Cyt b构建的最大似然树和遗传距离分析显示,巢鼠和红耳巢鼠有明显分化,两者之间的遗传距离达到了13%以上,同时各物种在不同地区间也存在一定的遗传分化。GenBank中两条产地为中国的巢鼠序列 (HM217360和HM217361) (Pagès et al., 2010),其与巢鼠的遗传距离为11.4% ~ 11.9%,与红耳巢鼠的遗传距离为13.7% ~ 13.8%,系统发育分析也显示其与巢鼠有较为明显的分化。这些结果提示我们巢鼠属仍可能存在未被认识的分类单元,巢鼠属分类研究仍需进一步加强。
参考文献
On the taxonomic status of the harvest mouse Micromys minutus (Rodentia: Muridae) from Vietnam
Report for October meeting, 1855
Micromys agilis, Kleinmaus, ein neues Säugthier der Fauna von Dresden
MEGA X: Molecular evolutionary genetics analysis across computing platforms
Revisiting the taxonomy of the rattini tribe: a phylogeny-based delimitation of species boundaries
Reisen durch verschiedene Provinzen des Russischen Reiches in den Jahren 1768-1774
New species and subspecies of mammals in Gaoligong Mountains (Ⅰ)
The origin of Swedish and Norwegian populations of the Eurasian harvest mouse (Micromys minutus)
A guide to the measurement of mammal skull Ⅲ: Rodentia and Lagomorpha
Phylogeographic patterning of mtDNA in the widely distributed harvest mouse (Micromys minutus) suggests dramatic cycles of range contraction and expansion during the mid- to late Pleistocene
PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies
高黎贡山的兽类新种和新亚种(一)
兽类头骨测量标准Ⅲ: 啮齿目、兔形目

 青公网安备 63010402000199号
青公网安备 63010402000199号{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}